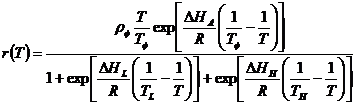|
The rice leaf folder, Cnaphalocrocis medinalis (Guenee) (Lepidoptera: Pyralidae) was earlier considered a minor pest in many Asian countries. However, it assumed major pest status since the 1980s causing serious economic loss in case of severe infestation coinciding with susceptible crop growth stages (maximum tillering to booting). The shift in pest status is mainly attributed to changes in crop management practices such as cultivation of high yielding varieties, adoption of higher plant density, planting either too early or too late, higher N fertilizer use and increased insecticide applications for pest control adversely affecting natural enemies of leaf folder. The pest undergoes two to three generations on rice crop. The first brood on rice is initiated mainly through immigrating adults while the second and third broods develop within the crop and are the most damaging . It is possible to forecast the timing of leaf folder activity in the field based on the concept of thermal time for completion of insect life cycle. To predict the timing of pest stages, user is required to provide inputs such as date of transplanting, current season weather data for the location, date of first observation of pest (BioFix Date) and pest stage observed on that date. Further user can select the applicable state/district.
Rice leaf folder species complex
Until 1981, Cnaphalocrocis medinalis Guenee was the only species considered as a rice leaf folder. The identification of Marasmia patnalis by Bradley in 1981, which caused leaf damage along with C. medinalis, led to the realization that there was a leaf folder complex. Four superficially similar species (C. medinalis, M. patnalis, M. exigua (Batler) and M. ruralis (Walker)) were recorded to be very common in rice growing areas [1]. In addition, M. bilinealis (Hampson), M. suspicalis (Walker), M. trapezalis (Guenee), and M. venilialis (Guenee) [2-3] were reported from South East Asian countries. Apart from these Brandia admixtalis (Walker), from Japan [4], and Brachmia aretraea Mayrick, a Gelichiid leaf folder from Indo-Malaya [5] were also reported as leaf folder pests of rice in India. However, C. medinalis and, M. patnalis are major widespread species [6-7]. Rice leaf folder insects are widely distributed in all the rice growing tracts of humid tropical and temperate countries in Asia, Oceania, Australia and Africa [6-11]. In India, leaf folder is either a regular or sporadic pest at most rice growing locations. Outbreaks of serious rice leaf folder infestations have been reported in many Asian countries, including China, India, Pakistan, Japan, Korea, Malaysia, Sri Lanka, and Vietnam [12-13]. In India, outbreaks of leaf folder on rice resulting in extensive damage were reported during 1977 in Uttar Pradesh [14], 1985 in Haryana [15] and 1998 in Tamil Nadu [16]. In 2012 kharif season, extensive damage due to leaf folder on Basmati rice varieties was observed in the month of October in Haryana and Punjab and on non-basmati rice varieties in Assam. Leaf folder outbreaks occur due to several factors. Reports mainly attribute to a reduced abundance of natural enemies either as a consequence of extreme weather conditions such as drought [17-19], or prevalence of conductive weather conditions 32-33°C temperature and relative humidity at early stage of the crop and rainfall or the indiscriminate use of broad-spectrum insecticides [20-21]. Often pest infestation is initiated and promoted on rice plants under shade found near field borders [16]. Leaf folder damage increases proportionately with increasing nitrogen fertilizer application [22]. Therefore, balanced fertilizer application is recommended to avoid outbreaks of C. medinalis [23-25]. Rice leaf folder is monophagous on rice (Oryza sativa) and undergoes complete development. However, insect life stages can partially develop on many host plants such as: Bracharia plantaginea B. mutica (Forssk.) Stapf Dactyloctenium aegyptium (L.) Willd Digitaria ciliaris (Retz.) Keel. Dinebra retroflexa Echinochloa colona (L.) Link E. crusgalli (L.) P. Beauv. spp. Hispidula (Retz.) Honda E. glabrescensMunro ex Hook. f. Eleusine indica (L.) Goertn. Leersia hexandra SW., Leptochloa chinensis (L.) Nees, Panicum repens L. Paspalum conjugatum Berg. P. distichum (L.) Willd., Setaria verticillata S. tomentosa Sorghum bicolor, Triticum aestivum Rice leaf folder development undergoes four life stages: egg, larva, pupa and adult. It requires 24 days for complete development of life cycle passing through five instars [40]. Adult Male and female moths are smaller in size and can be seen resting on walls near light sources after emergence. Males can be distinguished based on presence of dark brown androconial scales on midcostal vein of forewings. Adults live for about two weeks when provided with 10% sucrose and lay eggs in 2 to 3 days after emergence from pupa. Each mated female lays about 20 to 40 eggs per day with a lifetime fecundity of 300 eggs [26-28]. Female Male
Egg
Eggs are laid either singly or in groups arranged in longitudinal rows on the undersurface of the leaves, appear flat on the leaf surface and white in color. Eggs hatch in about 3 to 10 days depending on prevailing temperature and humidity. Larva Newly hatched larva is whitish yellow in color, body turns transparent green once the larva feeds on green matter. Larval stage undergoes 5 instars and lasts 13 to 37 days depending on temperature.
Pupa Final instar larvae spin silken webbing, either inside the leaf fold closed on both sides or behind a leaf sheath. Pupal period varies between 6 to 10 days depending on temperature.
Leaf folder damage can be observed at any stage of the crop growth but generally is conspicuous during active tillering to booting stage. Prior to feeding, larvae fold the leaves longitudinally by fastening the leaf margins with thread like silk. Contraction of silk strands results in a tubular structure open at both ends resulting in the leaf roll or leaf fold which is easily identified under field conditions. Sometimes a feeding chamber is created by stitching two or three adjacent leaves together [29]. Larvae feed inside the folds by scraping off the green mesophyll tissue between the veins. This results in leaves with linear, pale white striped injury symptom, while heavily injured leaves may appear papery white and dry up completely [30]. Feeding rate of the first three larval instars of C. medinalis on rice plants was significantly lower than the fourth and fifth instar larvae. Most often one larva is present in a folded leaf and after feeding for about 2-3 days, it moves on to another leaf. Thus each larva damages a number of leaves during its growth period [31]. Leaf damage results in a drastic reduction in photosynthetically active area which eventually affects plant growth and yield performance. Identifying key natural enemies and vulnerable life stages of rice leaf folder are essential for careful planning and developing practical management programs which intern helps in reducing the yield loss [39]. The pest has been recorded to cause 63% to 80% yield loss in rice [32-33]. At flowering stage, flag leaf area damage of above 25% resulted in more than 50% unfilled grains over control, indicating direct effect of yield reduction in rice [41]. Modeling development, survival and population increase Temperature is a key influencing factor on development of leaf folder populations under field conditions. To model the development, it is important to estimate lower and upper parameter thresholds for initiation and cessation of development, respectively. This is best done under constant temperatures under laboratory conditions. Complete development of individual rice leaf folder insects reared under a range of temperature conditions (18 to 32°C) was assessed keeping ecologically relevant relative humidity and photoperiod conditions constant across temperatures. The instar development durations were converted to developmental rates and subjected to linear and non-linear regression techniques. Linear models Two models were evaluated to estimate the linear relationship between ecologically relevant temperatures and the rate of development of the pest. The first was the thermal summation model [34] which is given by the expression,
Where, r is the rate of development (=1/ Development time (D) in days), T is ambient temperature (°C); intercept (a) and slope (b) are the model parameters. Thermal constant, k (= 1/b), is the number of degree-days (DDs) or heat units above the threshold needed for completion of an instar. Lower temperature threshold (Tmin) was determined as the x-intercept (= - a/b) which is the estimated lower temperature at which the rate of development is either zero or no measurable development occurs. The second linear model by Ikemoto and Takai [35] is given as,
Where, DT is theproduct of the duration of development, D (days), and temperature, T (°C), k is thermal constant and Tmin is the lower developmental threshold. Nonlinear models Two empirical nonlinear models were fitted to the instar specific developmental rate data to estimate the optimum temperature threshold (Topt) and upper temperature threshold (Tmax). Topt is the threshold temperature at which developmental rate is maximal, while Tmax is the lethal threshold at which development ceases. Lactin-2 model [36], Briere-1 model [37] were applied to assess the nonlinear relationship.In addition, a thermodynamic model referred to as the Sharpe - Schoolfield - Ikemoto model (SSI model) was used to estimate intrinsic optimum temperature for the pest [38]. All the nonlinear models described the relationship between developmental rate (1/D) and temperature (T). The Lactin-2 model [36] is given by the expression,
Where, D is the mean development duration in days, ρ is the composite value for critical enzyme-catalyzed biochemical reactionsas T increases to Topt, Δ is the difference between Topt and Tmax when thermal breakdown becomes the overriding influence and λ is a fitted coefficient that forces the nonlinear curve to intersect the x-axis and allows the estimation of lower developmental threshold. The Briere-1 model [37] is given by the expression,
Where, r is the developmental rate as a function of temperature (T), and ‘a’ is an empirical constant. The nonlinear thermodynamic SSI model (Shi et al., 2011) is given by the expression:
where, developmental rate (r) is a function of temperature, T (in absolute temperature, K) (273.15K = 0°C), R is the gas constant (1.987 cal/deg/mol), Δ HA is the enthalpy of activation of the reaction that is catalyzed by the enzyme (cal/mol), ΔHL is the change in enthalpy associated with low-temperature inactivation of the enzyme (cal/mol), ΔHH is the change in enthalpy associated with high-temperature inactivation of the enzyme (cal/mol), TL is the temperature at which the enzyme is half active and half low-temperature inactive (K), TH s the temperature at which the enzyme is half active and half high-temperature inactive (K), TΦ is the intrinsic optimum temperature at which the probability of enzyme being in the active state is maximal (K), and ρΦ is the mean development rate at the intrinsic optimum temperature (TΦ ) assuming no enzyme inactivation (day-1). For a complete description of the phenology model for leaf folder please refer Padmavathi et al., 2013 [42]. Survival and fecundity rates are used in construction of life tables to predict the potential population size and trend index under input weather conditions at the location of interest. 25 and 30°C were favourable temperature for survival and multiplication of the pest [42]. Estimated lower and upper thresholds and thermal constants (degree-days) from models were used to predict the percent development and timing of different life stages and cumulative development of immatures from egg to adult and generation based on observed stages under field conditions and input temperature data for the location. Predictions help in initiating field scouting, planning for ecofriendly interventions and timing of least disruptive insecticides managing pest infestations.




|